The Nuclear Envelope and Traffic between the Nucleus and the Cytoplasm
처음에는 핵의 존재 유무로 원핵세포와 진핵세포 구분 > 지금은 internal membrane system의 유무로 구분
The Nucleus
핵 존재의 특별성
; 유전정보를 가지고 있는 DNA를 막 안에 보호
; 유전물질의 공간적 구분/물질적 구분 이상으로, 유전자의 발현 조절 / 기능 조절의 새로운 차원
; 핵 내/외부의 물질 조성을 구분짓는 selective traffic(transport) – nuclear pore complex (특히 macromolecule의 수송에 있어서)
핵 구성체
; 2개의 막 (inner membrane/outer membrane)
; inner membrane 안쪽의 nulcear lamina
; nuclear pore complexes
; outer membrane은 ER membrane과 continuous, 일정 부분 공유
; inner / outer membrane 사이 공간은 perinuclear space; ER lumen과 연결 (서로 물질 왔다갔다 가능)
; inner membrane 안쪽에 선형단백질들이 복잡한 meshwork를 형성하여 단단한 구조 > nucleus membrane system이 일정한 모양을 유지할 수 있도록 지지 (cytoskeleton), 지지뿐 아니라 유전자 발현 조절 + chromatin 구조 조절에 있어서도 역할
전자현미경 이미지를 얻을 때 사용하는 테크닉
; 저온의 액체질소에 샘플을 넣은 후 깨기 > pore complex가 쉽게 보임 (even하기보다는 cluster되어 있는 것 관찰 가능)
Lamins
; 형태에 따라 단백질 구분했을 때(globular protein / fibrous protein) fibrous protein의 대표인 lamin 단백질
; lamin = globular domain + 이어주는 fibrous domain
; head-tail 만나서 dimer 형성, α-helix 부분이 coiled coil interaction (안정적)
; tetramer 형성; staggered array (한 dimer의 중간에 또 다른 dimer가 끼어 들어감)
; head-to-tail assocation; tetramer끼리 만나서 protofilament 형성(최종 완성)
; lamin을 포함하여 대부분의 intermediate filament들이 이런 식으로 protofilament 형성
; 이런 filament들이 인지질들에 붙어 nuclear skeleton 역할을 해주는 것, 그 전체 구조를 nuclear lamina라 부름
Lamins
; LBR(lamin B receptor) / emerin(nuclear envelope 내에 embedding되어 있는 단백질들)과 연결; strong interaction
; inner/outer membrane을 spanning하는 LINC complex; 안쪽으로는 lamina / 바깥쪽으로는 cytoskeleton과 interaction
; chromatin과도 강하게 association하고 있음
nuclear lamina의 구조 영향을 줄 수 있는 mutations
; lamin 자체의 mutation으로 인해 lamina가 제대로 형성되지 않는 경우
; emerin/LBR 등의 mutation으로 lamina가 핵막과 interaction을 잘 하지 못하는 경우
; nuclear skeleton이 잘 기능하지 못하는 현상 > 그런 경우 근육이 퇴화되는 질병 생기기 쉬움(nuclear skeleton/cytoskeleton mutation)
Nuclear pore complexes
; 거대한 구조, 구성 단백질들도 많음 최소 30개 이상, 구성에 참여하는 단백질들을 nucleoporins라 부름
; small polar molecule(hydrophobic region/hydrophilic region의 구분이 뚜렷한 물질들), ions(charged), macromolecule들의 traffic에 중요, cf. lipid-soluble한 것들은 인지질 2중막을 통해 쉽게 통과
; size에 따라 다르게 통과; 작은 애들은 free하게 통과, free diffusion(농도구배), 큰 애들은 여러 도움을 받아 통과
; eightfold symmetry 구조를 형성하고 있음
; 바깥쪽/안쪽 ring을 형성하고 있음 (cytoplasmic ring / nuclear ring)
; 사이에 기둥처럼 spanning하고 있는 column subunit + 그 위에 ring 구조
; cytoplasm 쪽으로는 선형으로 뻗은 filament 구조 형성 / 핵 안쪽으로는 fibril들이 nulcear basket 구조 형성
Selective transport of proteins to and from the nucleus
핵 밖에서 핵 안으로 들어가려는 macromolecule의 인증 수단인 NLS
; NLS는 단백질 내 특별한 서열정보; basic amino acid (positive charge) – single stretch, 일부 basic amino acid가 class 형성 + 한참 떨어진 데에도 존재 (공간적 분리) – bipartite
; 이러한 서열정보를 가지고 있는 단백질들은(NLS, NES) nuclear transport receptors (importin/exportin)라는 도우미들을 통해 들어오거나 나감
; importin과 exportin이 기능을 발휘하는 데에는 small GTP-binding protein인 Ran 단백질이 중요 (GTP bound일 때 활성, GDP bound일 때 불활성)
; adapter karyopherin이라는 또 다른 도우미 단백질과 함께 기능하는 importin들도 있음, heterodimer를 이루어 하나의 importin처럼 기능
; importin/exportin 둘 다로 dual role 하는 애들도 있음
Karyopherins
; nuclear transport receptor 중 가장 큰 protein family로 아미노산 서열 정보가 유사한 protein family
; 공통적으로 핵 안으로 거대물질을 들여보내거나 핵 밖으로 내보냄
ex. importin; Karyopherin family 중 가장 처음 발견됨, basic amino acid로 이루어진 NLS를 가지는 물질을 핵 안으로
ex. snurportin과 crm1; 핵에서 만들어진 snRNAs들이 나와서(by crm1) 세포질에서 단백질들과 만나 snRNPs 형성 > 형성된 snRNP는 핵 안으로 다시 들어가(by snurportin) 핵 안에서 기능
ex. transportin; 세포질에서 만들어진 mRNA binding protein들을 핵 안으로
ex. transportin과 importin7, crm1; ribosomal protein encoding mRNA는 핵 안에서 만들어짐 > 세포질에서 ribosomal protein 합성 > 얘네는 다시 핵 안으로 들어가서(by transportin/importin7) ribosome을 구성하는 40S, 60S subunit으로 ribosome assembly (at nucleolus) > 만들어진 40S, 60S subunit은 다시 핵 밖으로 나와서(by crm1) protein의 합성에 쓰임
ex. crm1; exportin 중 가장 흔함, leucine-rich sequence인 NES를 가지는 단백질들을 핵 밖으로, cytoplasm에서 기능하는 snurportin을 핵 밖으로
ex. exprotin-t; 세포질에서 기능하는 tRNAs를 핵 밖으로
ex. exportin5; 핵에서 만들어진 pre-miRNA > drosha에 의해 잘려서 pri-miRNA > 세포질로 나와서(by exportin5) dicer에 의해 잘리며 mature miRNA
general protein import 과정 (importin이 NLS을 가진 단백질을 핵 안으로 들여오는 과정)
; 세포질에서 핵 안으로 들어가야 하는 cargo protein의 NLS는 importin에 의해 인식됨
; cargo protein/importin complex 형성
; importin의 guide로 nucleus pore complex로 이동 및 통과
; 통과한 후 분리, cargo는 nucleus 내에 release, importin은 recycle되어 다시 밖으로 나가야 함
; Importin은 Ran/GTP에 의해 인식되어 complex를 이뤄 같이 나감
; 나가는 중에 cytoplasm filament에 anchor되어 있는 Ran GAP 만남 > Ran이 가지는 GTP 가수분해 활성을 촉진
; 나가며 Ran/GDP 형태가 되면 더 이상 interaction하지 못하니 cytoplasm에 importin이 release, 나온 Ran/GDP는 다시 핵 안으로 들어가야 함
; Ran/GDP를 위해 특수화된 importin인 NTF2가 binding하여 Ran/GDP를 import시켜줌
; 들어온 Ran/GDP는 Ran GEF를 만나 inactive한 Ran/GDP에서 다시 active한 GTP binding form이 됨
정리
; general importin의 도움을 받아 특수한 단백질이 핵 안으로 들어올 수 있음
; importin을 recycle하는 데 필요한 것이 Ran/GTP
; Ran/GTP를 다시 recycle하는 데 필요한 특별한 importin이 NTF2
; Ran GEF는 이 process에 있어 active한 Ran/GTP를 계속 생성
; importin과 결합한 Ran/GTP의 일방향적 transport를 가능하게 하는 게 Ran GAP, 얘가 cytoplasmic face에만 있기 때문(가수분해되어 세포질로 나간 Ran/GDP가 다시 importin을 데리고 들어오지 못하게 함)
general protein export 과정 (exportin이 NES를 가진 단백질을 핵 밖으로 내보내는 과정)
; NES를 가지고 있는 cargo protein이 핵 안에 존재
; exportin이 NES를 인식, exportin/cargo protein complex에 Ran/GTP가 binding하여 핵 밖으로 나감
; 내보내면서 Ran GAP이 GTP 가수분해, reversive process 불가능
; cargo protein이 release
; exportin은 karyopherin family를 위해 특화된 NTF2에 잡혀서 Ran/GDP까지 다 같이 complex 형성
; Ran/GDP를 recycle할 때 exportin도 같이 recycle되어서 들어옴
; Ran/GDP의 reactivation은 Ran GEF에 의해서
Transport of RNAs
핵 안에서 만들어져서 핵 밖에서 기능하는 RNA도 transport가 중요함
; RNA는 혼자 transport되지 않는다, protein과 complex를 이루는 것이(RNP complex 형성) 필수적
; tRNAs, rRNAs, miRNAs는 특화된 karyopherin family의 exportin이 존재한다 ex. Exportin-t, Exportin5
; mRNA는 exportin/karyopherin family dependent하게 transport가 이루어지지 않고, 대신 구분되는 exporter complex를 형성하여 이를 통해 핵공 밖으로 나간다, cytoplasmic face에만 존재하는 helicase에 의해 일방향적 transport가 진행된다
mRNA transport
; 핵 안에서 mRNA가 만들어짐. 5’cap과 poly A tail까지 모두.
; 만들어진 mRNA에 단백질들이 잔뜩 붙어 RNP complex 형성
; 붙은 단백질들 중 몇몇이 exporter complex 형성에 핵심적 역할을 수행하여 mRNA의 export를 가능하게 함, 그리고 이때 karyopherin family가 관여하지 않음
; 이 exporter complex는 exportin과 비슷하게 기능하면서 mRNA를 바깥으로 내보냄
; 나갈 때 helicase가 mRNA의 2/3차 구조를 바꿔주면서(stem 구조를 풀어주거나 만들어주면서) exporter complex를 형성하던 단백질들이 떨어져 나옴
; 다른 종류의 RNP complex가 세포질에 존재하게 됨
; exporter complex가 나갈 때에만 helicase가 작용하기 때문에 일방향적으로 진행됨, 바깥쪽에서는 exporter complex가 형성된다 하더라도 안으로 들어올 수 없음
=> mRNA는 다른 종류의 export complex를 형성하면서 나간다
snoRNA transport
; 핵의 인에서 기능하는 snoRNA(small nucleolar RNA)는 ribosomal RNA와 ribosomal protein간의 complex 형성 도움, ribosomal RNA의 final maturation도 도움
; ribosome biogenesis에서 중요한 역할
; snoRNA는 핵 안에서 만들어져서 핵을 떠날 필요가 없으니 계속 핵에 stay
snRNA transport
; snRNA는 혼자 기능하지 못하고 protein과 complex를 이뤄서 핵에서 기능해야 함
; snRNP complex를 만들기 위해 핵 밖으로 감 by 특화된 exportin인 Crm1(snRNA의 5’cap 인식)
; snRNP 형성 (functional assembly, 이때 기능을 취득)
; snRNP가 핵 안에서 기능하기 위해 들어옴 by 특화된 importin인 Snurportin
핵막을 경계로 하여 유전자 발현을 조절하는 mechanism의 또다른 예시들
• 염증반응에 관여하는 NF-kB, transcription factor
; NF-kB는 핵 안으로 들어와서 transcription factor로 기능해야 함, NLS를 가지고 있음
; 정상적으로는 NLS를 통해 importin과 binding하여 핵 안으로 들어옴
; 이 proces를 조절하는 새로운 메커니즘인 IkB
; IkB는 NLS를 통해 NF-kB와 interaction, complex를 이루면 NLS가 숨겨져서 importin에 의해 이동 불가능
; 대기하고 있다가 NF-kB가 사용되어야 하는 신호가 주어지면 IkB가 phosphorylation되면서 분해됨 > NLS의 노출
; NF-kB를 새로 만들지 않고도 필요시 IkB를 없앰으로써 빠르게 transcription factor의 기능을 발휘할 수 있게 함
; transcription factor를 이미 다 가지고 사용하지만 않고 있다가 사용할 필요성이 생기면 inhibitor를 빠르게 없애서 바로 사용
• Pho4, transcription factor
; Pho4의 NLS 부분이 phosphorylation되어 있으면 importin이 인식할 수 없음 (평소에)
; 필요한 신호가 오면 dephosphorylation시켜서(by phosphatase) NLS가 importin에 의해 인식되어 핵 안으로 들어가서 사용됨
The Organization of Chromatin
chromatin은 세포가 분열하면 compact해짐 > 짧고 굵은 모양으로 metaphase에 chromosome 형성
cf. 분열하지 않는 interphase에는 긴 상태로 존재
chromosome territory
; interphase 때 chromosome이라 할지라도 핵 내에 random하게 아무데나 존재하는 것이 아니다
; 핵 내에서 chromosome들이 왔다갔다 움직이는 상태가 아니다
chromosome 별로 각자의 방이 있다, 핵 내 공간 내 각 chromosome이 각자 안정적으로 있는 위치가 정해져 있다 (distinct region)
chromosome을 기반으로 일어나는 복제/전사는 핵 내에서 상당히 정해진 위치에서 일어난다
; replication, trasncription이 활발히 일어나는 region이 정해져 있다 (clustered regions)
핵 내 chromosome들이 특정한 위치를 점유하고 있다는 생각 자체는 아주 오래됨 since 1885, 구체적 컨셉으로 발달한 건 비교적 최근
아주 최근 개발된 연구기법인 3C
; 내가 원하는 유전자/chromosome의 특정한 부위가 핵 내 어떤 3차원적 공간에 존재하는지 mapping
; chromosome, 유전자 수준에서 3차원 시공간화, 특히 공간화
; chromosome territory 개념 하에서 가능한 기법 (random하게 위치하고 이동하는 거라면 할 필요 없음)
heterochromation과 euchromatin
; 응축되어 고밀도로 존재하는 부분 / 느슨한 형태로 존재하는 부분
; 실제 사용은 거의 되지 않는 부분 / 쉽게 발현되는 부분
; dense staining, 어두운 색 / 밝은 색
; nuclear envelope, nucelolus 부분 주위로 / 핵 기질에 넓게 퍼져 있음
; 이 개념은 epigenetics의 기반이 됨 > 유전자를 가지고 있더라도 둘 중 어떤 상태로 있는지가 중요, 표현형으로 연결될 수도 / mendel genetics에서 유전자를 가지고 있지 않은 것과 같은 결과가 나타날 수도
heterochromatin 다시 두 종류로
• constitutive heterochromatin
; 태어나서 죽을 때까지 계속 heterochromatin로만 남아 있음
; 유전자가 별로 없고 있더라도 거의 발현되지 않는 유전자들만 가지고 있음
; centromere, telomere, transposable element가 많이 존재하는 chromosome, …
• facultative heterochromatin
; 어떤 때에는 euchromatin로 바뀌어 있기도 하고, 세포종류마다/조절신호에 따라/외부 환경에 따라 hetero/eu를 왔다갔다 > 바뀌면서 위치 이동 (위치 이동하지 않는 예외도 있음)
; 생물학적 조절 측면에서 중요
; 마찬가지로 euchromatin도 heterochromatin으로 될 수 있음
TADs
; 한 chromosome 중에서도 특정하게 다른 부위와 구별될 수 있는 특정 chromosome 부위(한 chromosome 내에 여러 TAD 존재)
; 구분되는, 다른 chromosome인데도 둘 사이에 같은 region에 존재하는 부위
; 공간적으로 구분되는 domain, 한 chromosome의 특정한 segment 존재 / 서로 다른 여러 chromosome의 segment들이 공존
; 핵을 3차원적 구조로 바라보며 등장한 개념, 3C를 통해 다 발견할 수 있음
TAD 개념 하에 LAD/NAD가 있음
; LAD는 TAD 중 핵막 그 중에서도 nuclear lamina 근처에 있는 chromosome domain (핵막 근처 까맣게 staining 되는 게 LAD)
; NAD는 nucleolus 근처에 있는 chromosome domain (인 근처 까맣게 staining 되는 게 NAD)
; LAD나 NAD는 주로 heterochromatin 형성(인 가장자리, 핵막 가장자리)
chromosome의 특정한 부분은 얽히고 설키며 독립적인 3차원적 구조와 공간을 형성; TAD
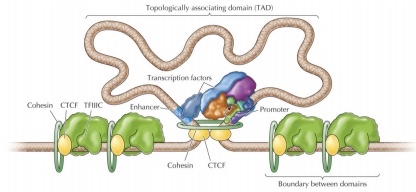
이 그림은 한 chromosome의 한 segement가 3차원적으로 독립된 구조를 형성한 경우
그러나 한 TAD 안에는 한 chromosome의 한 segment만 있는 것이 아니며 실제로는 인접한 다른 chromosome의 다른 segment도 함께 있을 수 있다
chromosome의 특정 위치 점유 확인
; 유전자 밀도가 높고 활발히 발현되는 chromosome 19, 핵 중심부에 치우쳐 있음
; 유전자 밀도가 낮고 유전자 발현이 적은 chromosome 18, 핵막 근처/핵 바깥쪽으로 치우쳐 있음
; 핵막 근처에는 주로 발현이 활발히 일어나지 않는 chromosome 부위가 위치해 있다
; 한 chromosome 중에서도 좀 더 활발히 발현되는 chromosome 부분은 핵막보다는 핵 안쪽으로 존재한다
; 각 chromosome들은 LAD/NAD association되어 있으면서도 핵에 나름 distinct region에 구분되어 존재한다
Nuclear Bodies
핵에서 일어지는 중심 metaboslim인 복제, 전사가 핵 모든 부분에서 random하게 even하게 진행되지 않음, 특정 부분에 집중해서 일어남
Replication factory
; DNA replication이 많이 일어나는 부위(hotspot)
; DNA replication 확인하는 방법 두 가지; BrdU / PCNA
• BrdU; DNA replication할 때 substrate로 incorporation됨(T 대신 들어가서) > florescent detection 가능 > 한 핵 내에서 일어나는 DNA replication activity 간접적으로 측정 가능
• PCNA; DNA replication을 담당하는 효소 complex 중 하나, labeling 해서 얘가 어디 많이 존재하는지 보는 것
Transcription factory
; replication factory 개념과 거의 똑같음
; RNA polymerase/특정 transcription factor를 추적해 봤더니 특정한 hotspot 내 집중해서 고밀도로 존재
; 고정된 DNA에 pol과 tf가 찾아와서 시작하는 줄 알았더니, 알고 보니 이미 pol과 tf가 transcription factory로 특정 부분을 점유하고 있고 그 내로 DNA가 들어갔다 나왔다 하는 것
; 한 transcription factory는 여러 chromosome에 의해, 한 chromosome의 여러 다른 region에 의해 동시에 공유될 수 있다
; replication과 transcription은 둘 다 이렇게 진행됨 (이해의 전환, DNA가 왔다갔다)
Nuclear body
; 핵 내 존재하는 특정한 구분되는 부분, 구획화되어 있는 부분 (replication factor/transcription factor도 일종의 nuclear body라 할 수 있긴 하나 그렇게 사용하지는 않음)
; 인지질 2중막으로 둘러싸여 있지 않음, 특정 protein/RNA가 고농도로 존재하면서 구획화
; lncRNA가 초기 nulcear body를 형성하는 데 중요한 역할을 한다(여러 molecule 모으는 scaffold 역할) + nuclear body는 일종의 droplet과 같은 생화학적 특성을 가진다
; nucleolus, cajal body, speckle(replcation/trasncription factory와 비슷), PML body, polycomb body
Nucleolus
; rRNA가 처음 trasncription되는 곳, rRNA가 mature되는 곳, ribosome assembly를 통한 ribosome subunit이 생성되는 곳
; rRNA 중 5.8S, 18S, 28S는 single unit으로 transcription되어(45S, by RNA pol 1) processing을 통해 세 개로 쪼개짐 cf. 대부분의 mRNA는 RNA pol 2에 의해 생성
; rRNA 중 5S는 별도로 유전자가 존재, RNA pol 3에 의해 transcription cf. RNA pol 3는 tRNA의 전사도 맡음
; 매우 많은 rRNA가 만들어져야 함, 따라서 매우 많은 rRNA copy를 동시에 가지고 있음. 분열을 활발히 하는 세포일수록
; rRNA 유전자 자체를 많은 copy로 보유하고 있음; 45S는 200개, 5S는 2000개 이상의 유전자 copy. 자기들끼리 한 region에 cluster된 걸 tandem array라 부름
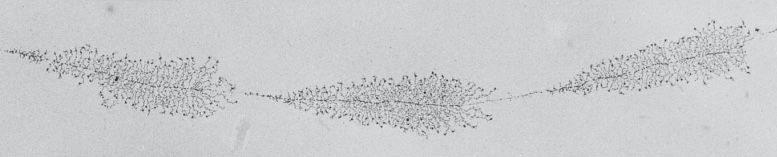
45S transcription unit 내에서도 많은 pol 1이 동시에 일하면서 긴 45S, 짧은 45S …
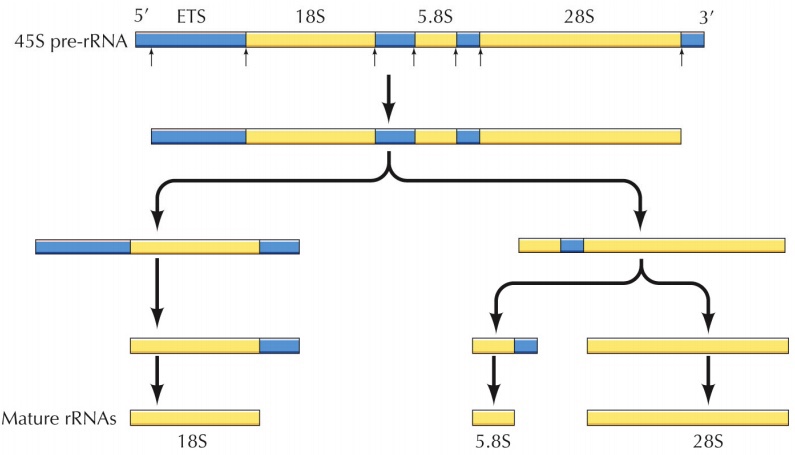
여러 processing 과정 거침 > mature rRNAs
; mature 18S (40S) / mature 5.8S + 28S (5S까지 합쳐서 60S)
; 5S rRNA는 별도 장소에서 transcription, 인으로 사후 투입, 다 만나서 60S subunit
; ribosomal protein들이 인으로 들어와서 다 같이 인에서 합쳐짐, subunit 형성, 핵 밖으로 나와서 protein synthesis factory 역할
snoRNA
; pre-rRNA는 maturation 될 때 잘리는 것뿐 아니라 특정한 부분이 methylation, uridine to pseudouridine 등 chemical modification 거침
; chemical modification 과정에서 snoRNA가 인에서 역할
; snoRNA는 인에 존재하는 protein들과 결합하여 snoRNP 형성
; snoRNP에서 snoRNA는 pre-rRNA와 base pairing (일종의 guide RNA 역할 > rRNA의 특정한 부분에 chemical modificaiton guide)
; RNP complex 내의 methylation시키는 효소를 통해(ex. fibrillarin) pre-rRNA의 methylation
*fibrillarin; RNA met 효소, snoRNP, cajal body, nucleolus에 존재
polycomb body
; polycomb protein complex(PRC), Prc1/Prc2 존재
; Prc2가 H3K27을 met (writer)
; Prc1은 그것을 인식, reading (reader)
; met 많이 된 euchromatin 부분은 facultative heterochromatin이 됨, 더 이상 사용 x (이런 eu>hetero의 변화도 핵 내 특정 부분에서만 일어나는 것)
; 이런 일을 수행하는 polycomb protein들도 핵 내에 특정 부위에 고밀도로 모여 있음(replication factory처럼)
cajal body
; 인에 비해 작음. cajal body에만 존재하는 coilin 단백질을 추적하면 확인 가능
; snRNA에 protein을 붙여줘서 snRNP의 final maturation 도움 (functional assembly된 snRNP가 오는 곳이 cajal body, 마지막 추가 protein assembly)
; 다른 종류의 RNP complex 형성도 이루어짐 ex. miRNA+dicer/AGO2 (RISC)
; snRNA의 chemical modification (by fibrillarin)
; cajal body에만 존재하는 RNA – scaRNAs
; snoRNA가 인에서 했던 일을 비슷하게 수행(guide RNA 역할 > snRNA의 chemical modification)
; teolmerase complex(RNP complex 중 하나)의 assembly
; 각 chromosome의 끝부분인 telomere에 simple sequence를 붙이는 telomerase, telomerase complex가 RNP complex 형태로 maturation 되는 장소
; 인과 비슷한 특성들 – RNA processing, assembly, modification
speckles
; snRNP가 assembly, maturation되면 snRNP 내의 snRNA가 하는 중요한 역할 중 하나인 mRNA splicing
; spliceosome complex 내에서 역할 수행
; cajal body에서 maturation된 snRNP는 speckle(splicing factor가 모여 있는 mRNA splicing factory)로 이동하여 기능
